Elastina es una proteína estructural que participa en la formación del tejido conectivo de los pulmones, los vasos sanguíneos y la piel. A diferencia del colágeno, que también se encuentra en el tejido conectivo, es muy elástico. Las moléculas de elastina se interconectan entre sí en el área extracelular.
¿Qué es la elastina?
Todos los vertebrados contienen elastina, la proteína de fibra. Es una proteína estructural que se encarga de dar forma a órganos tan importantes como los pulmones, los vasos sanguíneos o la piel. Junto con el colágeno, forma el tejido conectivo de estos órganos.
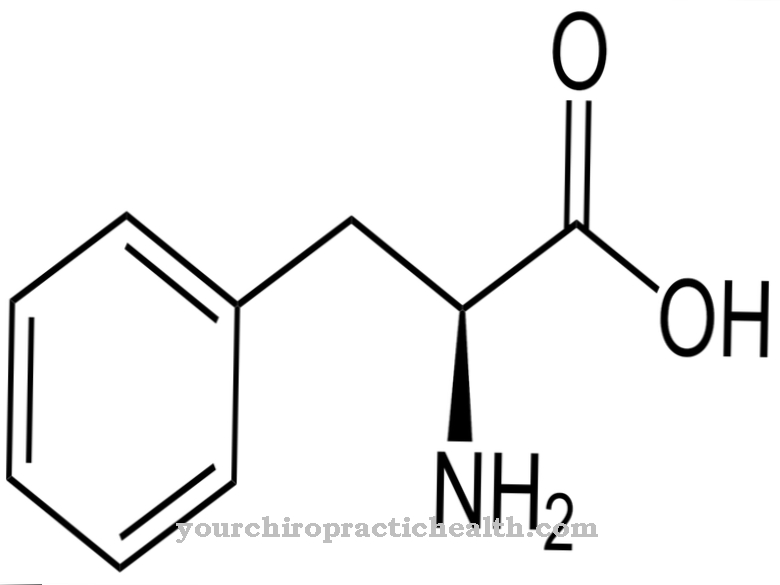
Las propiedades de la elastina y el colágeno se complementan. La elastina, como su nombre indica, es muy elástica en contraste con el colágeno. Esto hace que el tejido conectivo de la piel, los pulmones y los vasos sanguíneos sea elástico y maleable. Las funciones de estos tres órganos requieren un cambio de tamaño constante. La elastina se compone principalmente de los aminoácidos alanina, glicina, prolina, valina, lisina, leucina e isoleucina. Las áreas hidrofóbicas e hidrofílicas se alternan dentro de la molécula.
Las unidades características de los cuatro aminoácidos alanina, prolina, glicina y valina se repiten en cada dominio hidrófobo. Las zonas hidrofílicas contienen principalmente lisina. El residuo de lisina se oxida a alisina por la enzima lisil oxidasa. El grupo amino terminal se reemplaza por un grupo carboxilo. Los residuos de lisina de las diversas cadenas de proteínas se combinan entre sí para formar una desmosina en forma de anillo y, por tanto, entrecruzan las diferentes cadenas entre sí.
Función, efecto y tareas
Como proteína estructural dentro del tejido conectivo, la elastina tiene la función de asegurar la forma y elasticidad de los pulmones, los vasos sanguíneos y la piel. Los tres órganos dependen de la flexibilidad del tejido conectivo. Están sujetos a constantes cambios de volumen.
Como proteína estructural, el tejido conectivo tiene principalmente colágeno. Es resistente al desgarro, pero sería demasiado rígido como único elemento estructural. Solo la combinación de las propiedades de la elastina y el colágeno permite que el tejido conectivo se vuelva elástico y al mismo tiempo resistente al desgarro. El componente básico de la elastina es la tropoelastina. La tropoelastina se compone de dominios hidrófobos e hidrófilos alternados. Tiene una masa molecular aproximada de 72 kilodaltons. Las unidades de tropoelastina se interconectan entre sí en los residuos de lisina.
Si bien la tropoelastina es soluble en agua debido a sus muchos dominios hidrófilos, se niega la solubilidad en agua del polímero reticulado. La tropoelastina se forma dentro de las células y llega al área extracelular a través del transporte de membrana. Aquí es donde tiene lugar la interconexión de los bloques de construcción básicos, formándose unidades de desmosina en forma de anillo en los puntos de interconexión. Tres residuos de alisina y un residuo de lisina siempre participan en la formación de desmosina. Dado que la alisina es un producto de oxidación de la lisina, en última instancia, cuatro residuos de lisina se unen entre sí.
Esta forma de conexión confiere a la elastina su elasticidad especial. La reticulación también protege a la elastina de la desnaturalización y degradación por casi todas las proteasas. Sin embargo, la enzima elastasa es una excepción: es la única proteasa capaz de degradar la elastina. De esta forma se descomponen las elastinas ingeridas a través de los alimentos.
Educación, ocurrencia, propiedades y valores óptimos
Como ya se mencionó, la elastina es un componente necesario del tejido conectivo de los pulmones, los vasos sanguíneos y la piel. Esto afecta a todos los vertebrados. El bloque de construcción básico tropoelastina apenas se puede detectar en el tejido animal. Después de la conversión de los residuos de lisina en alisina por la lisil oxidasa, tres residuos de alisina se entrecruzan inmediatamente con un residuo de lisina. La elastina se produce casi exclusivamente en su forma de red.
Sin embargo, la detección de tropoelastina en experimentos con animales al inhibir la síntesis de lisil oxidasa fue exitosa. Si falta esta enzima, no hay conversión de lisina en alisina y, por tanto, tampoco formación de elastina. Debido a la resistencia de la elastina a la degradación por proteasas, la piel, los pulmones y los vasos sanguíneos están idealmente protegidos. El efecto degradante de la elastasa está limitado por los inhibidores de elastasa.
Enfermedades y trastornos
Las mutaciones en el gen ELN pueden causar enfermedades hereditarias en las que se modifica la estructura de la elastina. En la llamada dermatocalasia, hay cambios en el tejido conectivo, que se manifiestan en una piel flácida e inelástica que se hunde en pliegues.
La enfermedad puede ser tanto adquirida como hereditaria. Se observan agrupaciones familiares. Además de muchos otros síntomas, esta debilidad del tejido conectivo también ocurre en el síndrome de Williams-Beuren. Esta es también una anomalía estructural hereditaria de la elastina. La causa de esta enfermedad es una mutación en el cromosoma 7. Además, también existe una estenosis aórtica congénita, que se basa en una alteración en la estructura de la elastina. La arteria principal del corazón se estrecha. El flujo de sangre desde el ventrículo izquierdo al torrente sanguíneo se retrasa.
La insuficiencia cardíaca se produce a largo plazo. Del cinco al seis por ciento de todos los defectos cardíacos congénitos son estenosis aórticas congénitas. También se supone que algunas formas del síndrome de Ehlers-Danlos son malformaciones de elastina. Esta enfermedad se caracteriza por una piel que se puede estirar demasiado, que se llama piel de goma. La debilidad del tejido conectivo afecta a muchos órganos, incluidos el corazón y el tracto digestivo. El síndrome generalmente se hereda como un rasgo autosómico dominante.
En el llamado síndrome de Menkes, además de muchos otros síntomas, también existe una debilidad del tejido conectivo, cuya causa se encuentra en una síntesis alterada de elastina. En realidad, el síndrome de Menkes se caracteriza por una interrupción en la absorción de cobre en el cuerpo. Sin embargo, el cobre es un cofactor de muchas enzimas. Entre otras cosas, esto también incluye lisil oxidasa. Sin cobre, la enzima es ineficaz. Ya no se produce la conversión de los residuos de lisina en alisina. Como resultado, la reticulación de los residuos de lisina a desmosina ya no puede funcionar.


.jpg)